ВВЕДЕНИЕ
ГЛАВА 1. ИСТОРИЯ ИЗУЧЕНИЯ КВОРУМ-ЗАВИСИМЫХ СИСТЕМ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНОВ БАКТЕРИЙ
или напишите нам прямо сейчас:
⚠️ Пожалуйста, пишите в MAX или заполните форму выше.
В России Telegram и WhatsApp блокируют - сообщения могут не дойти.
ГЛАВА 2. КВОРУМ-ЗАВИСИМЫЕ СИСТЕМЫ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ
2.1 Общая характеристика
2.2.. Системы типа LuxR/LuxI
2.3. АГЛ-зависимая система Vibrio harveyi
2.4 АИ-2 — системы
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ПРИЛОЖЕНИЕ
ВВЕДЕНИЕ
В естественных условиях бактерии обычно существуют в составе сложных прокариотических сообществ, где они находятся в постоянном взаимодействии — как конкурентном, так и кооперативном — с другими микроорганизмами. Жизнь в таких условиях способствовала возникновению специальных систем обмена информацией, которые бактерии используют для контактов с клетками своего вида, а также для межвидовых и даже междоменных взаимодействий. [Atkinson, Williams, с. 959].
Одним из наиболее изученных способов бактериального межклеточного сигналинга является чувство кворума (quorum-sensing, QS). Чувство кворума — это процесс межклеточной коммуникации, который обеспечивает скоординированную изменение экспрессии генов в ответ на изменения плотности бактериальной популяции. В основе такого типа регуляции лежит способность бактерий производить, секретировать и распознавать специальные сигнальные молекулы – аутоиндукторы. Концентрация аутоиндуктора, прямо зависящая от численности клеток в окружающей среде, воспринимается ими как сигнал к активации и/или супрессии экспрессии определенных генов [Papenfort, Bassler, с. 576].
Хотя чувство кворума обнаружено у самых разных групп бактерий, наиболее изученными являются кворум-зависимые системы грамотрицательных бактерий — обширной и разнородной группы прокариотических микроорганизмов, имеющих наружную и внутреннюю мембрану с периплазматическим пространством между ними, а также специфический химический состав клеточной стенки [Лысак, с. 43-44]. Показано, что у грамотрицательных бактерий кворум-зависимые системы участвуют в регуляции десятков самых разнообразных клеточных процессов, многие из которых имеют важное значение для человека. [Henke, Bassler, с. 648]. Например, под контролем чувства кворума находится секреция факторов вирулентности и способность к образованию биопленок такими клинически значимыми патогенами, как синегнойная палочка Pseudomonas aeruginosa [Schuster and Greenberg], холерный вибрион Vibrio cholerae [Higgins, Pomianek, Kraml] и др.
Данная работа посвящена обзору известных кворум-зависимых систем регуляции экспрессии генов грамотрицательных бактерий. Актуальность выбранной темы обуславливается тем, что исследования кворум-зависимых систем потенциально имеют большое прикладное значение, например, для лечения хронических бактериальных инфекций. В то же время исследования кворум-зависимых систем дают ответы на фундаментальные вопросы о природе и возможностях межклеточного сигналинга у прокариот. Объектом исследования является регуляция генной экспрессии грамотрицательных бактерий, а предметом исследования — кворум-зависимые системы регуляции микроорганизмов этого типа.
Цель настоящей работы состоит в анализе известных кворум-зависимых систем регуляции генной экспрессии грамотрицательных бактерий. Для достижения поставленной цели нами были сформулированы следующие задачи:
- Изучить научную литературу, посвященную кворум-зависимым системам регуляции генной экспрессии бактерий
- Рассмотреть историю изучения кворум-зависимых систем регуляции экспрессии и основные направления их исследований, актуальные на сегодняшний день
- Описать кворум-зависимые системы регуляции экспрессии грамотрицательных бактерий и выделить их характерные особенности
ГЛАВА 1. ИСТОРИЯ ИЗУЧЕНИЯ КВОРУМ-ЗАВИСИМЫХ СИСТЕМ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНОВ БАКТЕРИЙ
1.1 Основные этапы развития исследований чувства кворума
В истории изучения бактериального чувства кворума можно условно выделить три этапа. Первый этап, пришедшийся на 1960-1980 годы, является периодом становления данного направления исследований. Именно в эти годы было открыто и изучено само явление чувства кворума, а также показаны общие механизмы работы “каноничных” кворум-зависимых систем регуляции экспрессии.
Первым, кто заговорил о бактериальной коммуникации, был венгерский микробиолог А.Томаш, изучавший в 1960-х годах процесс приобретения компетентности бактериями Streptococcus pneumoniae. Он пытался объяснить необычную закономерность: количество компетентов в культуре возрастало по экспоненте, причем этот эффект демонстрировал зависимость только от концентрации клеток. Томаш использовал достаточно простые микробиологические методы: он помещал некомпетентные клетки в фильтрат культуральной жидкости компетентных клеток, наблюдая при этом увеличение скорости трансформации последних. Томаш предположил, что способность к приобретению компетентности регулируется неким химическим веществом, секретируемым S. pneumoniae в среду и действующим только на клетки данного вида. Томаш назвал данное химическое вещество «активатором«, выделил его из культуральной жидкости и показал, что в очищенном виде «активатор« индуцирует приобретение клетками компетентности. На основе этого Томаш заключил, что S. pneumoniae используют «активатор» для межклеточной коммуникации [Tomasz, с. 155-157].
Чуть позже, в конце 1960-х — начале 1970-х годов, в США сформировалась группа ученых, занимавшаяся работой люциферазы — фермента, обеспечивающего способность к биолюминесценции — морских бактерий рода Vibrio. Биолюминесценция в культуре клеток V. fischeri подчинялась таким же закономерностям, что и способность к компетентности клеток S. pneumoniae: в свежеинокулированных колбах сперва наблюдалась задержка эмиссии света, а затем, через какое-то время, свечение культуры резко возрастало. Исследователи Нильсон, Платт и Хастингс предположили, что задержка в свечении происходит из-за инактивации транскрипции гена люциферазы; при этом в ходе фазы роста культуры происходит активация некоего химического фактора, который и запускает быструю и преимущественную транскрипцию люциферазных генов. Это явление они назвали «аутоиндукцией», а искомый химический фактор — «аутоиндуктором». [Nealson, Platt, Hastings, 1970, с. 313-322]. Двумя годами позже, уже на мутантных штаммах V. fischeri, было показано, что данный аутоиндуктор синтезируется самими бактериями и является веществом с низкой молекулярной массой [Eberhard A., 1972].
Эти работы привлекли некоторый интерес научного сообщества, и в 1970-е годы появился ряд статей, посвященных явлению «автоиндукции» у морских люминесцирующих бактерий [Nealson, Hastings, 1979, с. 501-505]. В следующее десятилетие было сделано несколько важных открытий. Во-первых, была установлена химическая структура аутоиндуктора V. fischeri, а также исследованы биохимические пути его синтеза. Аутоиндуктором оказался (N-(-3-оксогексаноил)-L-гомосерин лактон — малая молекула с гомосеринлактоновым кольцом и боковой ацильной цепью [Eberhard et al. 1981, с. 2444]. Во-вторых, были определены гены, ответственные за биолюминесценцию (lux-гены) и выяснен общий механизм работы lux-оперонов. В частности, было показано, что продукты генов luxR и luxI регулируют транскрипцию lux-генов, причем белок luxR работает и как рецпетор для аутоиндуктора и, одновременно, как активатор транскрипции. Белок luxI же является ацилгомосеринлактон-синтазой, то есть ферментом, катализирующим реакцию синтеза аутоиндуктора. [Engebrecht, Nealson, Silverman, 1983; Engebrecht, Silverman, 1984].
С распространением технологии секвенирования, давшей в руки исследователей мощный инструмент сравнительного анализа генов и геномов связан второй этап в истории изучения чувства кворума — этап расширения исследовательского поля. В 1989 году была получена и опубликована полная нуклеотидная последовательность lux-регулона V. fischeri [Baldwin, Devine, Heckel, Lin, Shadel]. После этого стало появляться всё больше работ, показывающих, что системы аутоиндукции, похожие на LuxR/LuxI систему, функционируют и не только в Vibrio spp., но и в других грамотрицательных бактериях. Так, в начале 1990-х годах у Pseudomonas aeruginosa сперва был найден белок LasR, гомологичный LuxR и участвующий в регуляции синтеза эластазы — протеолитического фермента инвазии [Gambello, Iglewsk, с. 3000-3009], а затем и синтаза LasI, гомологичная LuxI и катализирующая продукцию видоспецифичного ацилгомосеринлактона [Passador, Cook, Gambello, Rust, Iglewski]. Примерно в то же время было показано наличие системы аутоиндукции у фитопатогенов Agrobacterium tumefaciens (функция — регуляция передачи Ti-плазмид во время конъюгации) и у Erwinia carotovora (регуляция синтеза факторов вирулентности). Тогда же, в 1994 году, впервые было использовано понятие «бактериального кворума» [Fuqua, Winans, Greenberg, c. 273].
В 1995 году были опубликованы сразу две работы, указывающие на наличие сигнальных молекул (или, как их назвали исследователи, «феромонов») пептидной природы у грамположительных патогенных бактерий Streptococcus pneumoniae [Håvarstein, Coomaraswamy, Morrison] и Staphylococcus aureus [Ji, Beavis, Novick]. Дальнейшее изучение этих систем показало, что кворум-зависимая регуляция экспрессии у грамположительных бактерий устроена несколько сложнее, чем у грамотрицательных. Во-первых, пептидные аутоиндукторы синтезируются в виде предшественников, которые затем подвергаются процессингу и переносятся из клетки во внешнюю среду специальными АТФ-связывающими кассетными транспортерами. Во-вторых, рецепция осуществляется, как правило, за счет работы двухкомпонентной системы, в которую входят сенсорная мембранносвязанная гистидин-киназа и белок — регулятор ответа. Связывание пептидного аутоиндуктора с киназой фосфорилирует белок-регулятор ответа, который запускает фосфорилирующий каскад, что в конечном итоге приводит к активации транскрипции таргетных генов [Kleerebezem et al, с. 896 ].
Таким образом, к середине 1990-х годов стало понятно, что кворум-зависимая регуляция характерна для внутривидовых взаимодействий как грамотрицательных, так и грамположительных бактерий. Параллельно с этим еще на заре исследований чувства кворума было сделано предположение о том, что существует некий универсальный аутоиндуктор, благодаря которому может происходить обмен сигналами между клетками разных видов. Данная гипотеза основывалась на данных исследования, где было показано, что более десятка морских грамотрицательных бактерий синтезируют химическое вещество, которое с той или иной эффективностью имитирует действие аутоиндуктора Vibrio harveyi и вызывает биолюминесценцию в культуре V.harveyi [Greenberg, Hastings, Ulitzur, с. 90].
Для проверки этого предположения требовалось уточнение механизмов работы lux-оперона V.harveyi. Оказалось, что, хотя структурные гены lux-оперона V.harveyi идентичны V. fischeri, расположение и организация генов-регуляторов lux-оперона V.harveyi сильно отличалось от каноничной АГЛ-зависимой модели, описанной для V. fischeri [Miyamoto, Boylan, Cragg, Meighen, с. 193]. У V. fischeri гены, кодирующие главные регуляторные белки luxR и LuxI, находятся в проксимальной области промотора, прилегающего к lux-оперону luxCDABEGH. Однако анализ данной области у V.harveyi не показал наличия в ней гомологов luxR и luxI генов. Более того, гены, гомологичные luxI и luxR, вообще не были найдены на хромосоме V. harveyi [Miyamoto, Graham, Meighen, с. 1559-1561]. При этом у V. harveyi был найден регуляторный ген-активатор транскрипции lux-оперона, также получивший название luxR, однако не имеющий гомологии с luxR геном V.fischeri. Более того, luxR активация транскрипции у V. harveyi не зависела от ключевого параметра — плотности клеточной культуры [Swartzman E., Silverman M., Meighen, с. 7490-7492]
Именно эти данные — способность клеток V. harveyi к восприятию аутоиндукторов других видов и отсутствие гомологов известных белков-регуляторов lux-оперона — привлекли внимание группы американских ученых под руководством Бонни Басслер. Используя методы транспозонного мутагенеза, они идентифицировали участки ДНК, функционально важные для биолюминесценции у мутантных штаммов V.harveyi с ослабленной работой lux-оперона, затем секвенировали данные участки и пытались “собрать” из них полноценно работающие регуляторы lux-оперона. Басслер с коллегами установили, что регуляторов lux-оперона у V.harveyi не два, как считалось ранее, а, как минимум, три — luxL, luxM (задействованы в синтезе аутоиндуктора), luxN (участвует в рецепции аутоиндуктора). При этом аминокислотная последовательность белков LuxL и LuxM не походила ни на один известный на то время белок, а вот белок LuxN содержал участки, схожие одновременно с последовательностями сенсорной гистидин-специфичной протеинкиназы и белка-регулятора ответа двухкомпонентных систем. [Bassler, Wright, Showalter, Silverman, с 781-783]. Годом позже этой же группой были установлены дополнительные белки-регуляторы LuxP, LuxQ и LuxO, а также предложена схема регуляции lux-оперона V.harveyi (см. Приложения, рисунок 1) в основе которой лежала идея о существовании двух связанных кворум-зависимых систем регуляции экспрессии, имеющих разные аутоиндукторы, но выполняющих общую функцию репрессии транскрипции lux-оперона. [Bassler, Wright, Silverman, с.281-283]. Вскоре был определен ген АГЛ-синтазы V. harveyi, получивший название luxS. Что интересно, ген luxS сразу был идентифицирован в нескольких неродственных видах бактерий [Surette, Miller, Bassler, с. 1639-1644], и довольно скоро было показано, что его гомологи обнаружились в более чем 500 бактериальных геномах [Pereira, Thompson, Xavier, с. 158].
Insight into AI-2 biosynthesis came 2 years later. Genome sequence analysis demonstrated that the Borrelia burgdorferi luxS gene was the third in a three-gene operon (Schauder et al., 2001; Winzer et al., 2002b). The two other genes, pfs and metK, encode enzymes involved in the activated methyl cycle (AMC), an important metabolic pathway for the recycling of S-adenosylmethionine (SAM), the major methyl donor in the cell (Miller & Duerre, 1968; Schauder et al., 2001). The release of the activated methyl group from SAM to an acceptor molecule gives rise to a toxic intermediate, S-adenosylhomocysteine (SAH), which is converted by Pfs to S-ribosylhomocysteine (SRH) (Fig. 1). Two independent groups showed that LuxS catalyzes the cleavage of SRH to homocysteine and a compound, at that time unidentified, with AI-2 activity (Schauder et al., 2001; Winzer et al., 2002b).
Идея о межвидовой бактериальной коммуникации быстро стала очень популярной среди исследователей, и были предложены первые подходы по её использованию в лечении инфекционных заболеваний:
1.2 Актуальные направления исследований чувства кворума
биосенсоры
На основе бактерий Escherichia coli сконструированы высокочувствительные специфические lux биосенсоры для детекции ионов металлов и металлоидов (ртуть, кадмий, серебро, медь, мышьяк, сурьма), а также алкилирующих агентов. Бактерии содержат гибридные плазмиды pArsR’::lux, pMerR’::lux, pCopA’::lux, pAlkA’::lux, в которых транскрипция геноврепортеров luxCDABE Photo rhabdus luminescens осуществляется с индуцируемых промоторов: arsоперона, merоперона, гена copA и гена alkA соответственно. Проведено измерение основных характеристик (пороговая чув ствительность, амплитуда и время ответа) luxбиосенсоров. Показано, что окисление 1,1диметил гидразина перекисью водорода приводит к появлению в клетке алкилирующих соединений, обла дающих сильным генотоксичным действием.
ГЛАВА 2. КВОРУМ-ЗАВИСИМЫЕ СИСТЕМЫ РЕГУЛЯЦИИ ЭКСПРЕССИИ ГРАМОТРИЦАТЕЛЬНЫХ БАКТЕРИЙ
2.1 Общая характеристика
Для грамотрицательных бактерий показано использование, как минимум, трех кворум-зависимых систем. Первая — система типа LuxR/LuxI. Для неё характерны следующие общие черты. Во-первых, аутоиндукторами (АИ) являются, как правило, ацил-гомосеринлактоны (АГЛ). Благодаря своей низкой молекулярной массе и отсутствию заряда они способны свободно диффундировать через бактериальную мембрану по градиенту концентрации. Во-вторых, рецепторы для аутоиндукторов располагаются либо в цитоплазме, либо на внутренней клеточной мембране. В-третьих, синтез аутоиндукторов грамотрицательных бактерий регулируется по типу положительной обратной связи (feed-forward loop), который позволяет бактериям лавинообразно увеличивать транскрипцию нужных генов при достижении пороговой численности [Papenfort, Bassler, с. 576].
Вторая система кворум-зависимой регуляции, найденная у грамотрицательных бактерий — это система АИ-1 у Vibrio harveyi. По своей структуре эта система больше напоминает грамположительные бактерии, но выполняет функции внутривидовой для V.harveyi
Третья система — АИ-2, чья роль неясна, но есть теории. гипотеза 1 — что это коммуникационная межвидовая система. Вторая гипотеза — это необходимо для environmental sensing, третья — это просто способ утилизации токсичных продуктов обмена
Рассмотрим данные системы.
2.2. Системы типа LuxR/LuxI
АГЛ-зависимая система (она же — система типа LuxI-LuxR), обнаруженная у бактерии Vibrio (Aliivibrio) fischeri, стала первой и потому классической моделью кворум-зависимой регуляции экспрессии грамотрицательных бактерий.
- fischeri — симбиотические бактерии, колонизирующие световые органы морских животных (в частности, осьминога Euprymna scolopes). Способность к биолюминесценции у V. fischeri обеспечивается действием комплекса ферментов — люциферазы, редуктазы жирных кислот и оксидоредуктазы, главным из которых является люцифераза, которая катализирует реакцию окисления жирного альдегида (RCHO) кислородом воздуха, причем в качестве кофактора выступает восстановленный флавинмононуклеотид (FMNH2). Реакция сопровождается излучением кванта света (λ=490 нм). Синтез жирного альдегида, являющегося субстратом для люциферазы, катализируется редуктазой жирных кислот. Наконец, оксидоредуктаза способствует реакции восстановления кофактора FMNH2 из окисленной формы [Engebrecht, Nealson, Silverman, с 774].
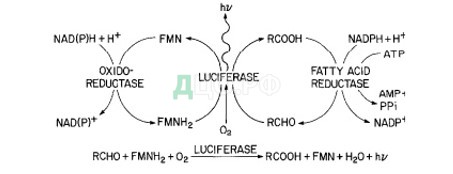
Схема химических реакций биолюминесценции
[Engebrecht,Nealson, Silverman, с 774]Гены, ответственные за биолюминесценцию у V.fischeri организованы в Lux-регулон, в состав которого входят два оперона: luxR и luxICDABEG, причем каждый из них находится под своим промотором. Гены luxA и luxB кодируют α и β субъединицы люциферазы. Продуктами генов luxC, luxD, luxE являются три субъединицы редуктазы жирных кислот. Наконец, luxG кодирует FMN-NADH оксидоредуктазу [Manukhov, Khrul’nova, Baranova, Zavilgelsky, с 3999].
Главными регуляторными компонентами Lux-регулона являются гены luxI и luxR. LuxI — ацилгомосеринлактон-синтаза (АГЛ-синтаза), фермент, катализирующий реакцию синтеза аутоиндуктора, которым у V.fischeri является N-(-3-оксогексаноил)-L-гомосерин лактон. Наиболее эффективными субстратами для LuxI являются интермедиаты пути биосинтеза жирных кислот, в частности, S-аденозилметионин (SAM) и ацил-ацил-переносящий белок (acyl–acyl carrier protein) [Schaefer et al., с 9508].
Второй ключевой регулятор — гетеродимерный белок LuxR, N-концевой домен которого имеет рецепторную функцию, а С-концевой — активаторную. Связывание N-концевого домена с аутоиндуктором приводит к конформационным перестройкам LuxR и модулирует активность С-концевого домена. С-концевой домен имеет ДНК-связывающий мотив спираль-поворот-спираль, благодаря которому он взаимодействует с lux-боксом — палиндромной последовательностью в 20 нуклеотидных пар (н.п.), расположенной на 42.5 н.п. выше точки старта транскрипции lux-оперона [Kolibachuk, Greenberg, с. 4127].
In LuxR, the N-terminal domain is believed to be membrane associated and to function as a receptor for the 3-oxo-C6-HSL ligand (15, 17). In response to binding the 3-oxo-C6-HSL, the N-terminal domain modulates the activity of the C-terminal domain (CTD). Binding of 3-oxo-C6-HSL permits multimerization of LuxR and subsequent activation of transcription of the lux operon, which is carried out by the CTD (3, 4). The CTD of LuxR has a helix-turn-helix motif and binds to a region of DNA termed the lux box, which is 20 bp long and has a dyad symmetry centered at a position −42.5 bp upstream of the transcription start site for the lux operon (5, 9). When LuxR functions as an activator of transcription at the luxI promoter, it is proposed to function as a homodimer in an ambidextrous manner similar to the manner observed for the cyclic AMP receptor protein at a class II-type promoter (1, 25) contacting both the alpha and sigma subunits of RNA polymerase (10, 16). Full-length LuxR protein was purified in the presence of 3-oxo-C6-HSL, which permitted its binding to the lux box to be demonstrated in vitro. Binding of 3-oxo-C6-HSL to LuxR appeared to be reversible in this system (34). In this study we examined the ability of LuxR to bind to the promoters of additional genes in V. fischeri MJ-100, leading to activation of transcription.
Структура lux-регулона V.fischeri
[Manukhov, Khrul’nova, Baranova, Zavilgelsky, с 3999]Из структуры lux-оперона V. fischeri видно, что LuxI — ген, кодирующий АГЛ-синтазу, также входит в его состав. Это обеспечивает регуляцию транскрипции по типу положительной обратной связи: связывание C-домена LuxR в комплексе с АГЛ с lux-боксом приводит к лавинообразному увеличению концентрации в среде всех продуктов, необходимых для проведения реакции биолюминесценции, что и объясняет экспоненциальный рост проявления данного свойства клеток V. fischeri. []
Белок LuxR
Синтезированные молекулы АГЛ свободно проходят через клеточную мембрану, двигаясь по градиенту концентрации, и оказываются во внешней среде [Kaplan, Greenberg, с 1213-1214]. С увеличением численности бактерий концентрация АГЛ в среде растет до достижения порогового уровня. После этого молекулы АГЛ начинают двигаться обратно в клетку, где связываются со вторым регулятором QS-системы — белком LuxR []
LasR/LasI у P. aeruginosa
работает по такому же типу. LuxI-гомолог LasI участвует в синтезе аутоиндуктора оксододеканоил-ГЛ. LuxRгомолог LasR и оксододеканоил-ГЛ необходимы для активации генов вирулентности, кодирующих эластазы (LasA и LasB), алкалинпротеазу (Apr), экзотоксин А (toxA). еще участвует в биопленках
RhlR/RhlI P. aeruginosa
состоит из активатора транскрипции RhlR и белка RhlI, ответственного за продукцию двух аутоиндукторов: доминирующего бутаноил-ГЛ и минорного гексаноилГЛ. реулирует продукцию рамнолипидов, эластазы, пиоционина и липазы
LuxR/I-type systems were identified in more than 70 Gram-negative species (Henke and Bassler, 2004a). Each LuxR-type sensor protein is very specific at detecting a particular AHL signaling molecule. The AHL-type pheromones differ in their acyl chain length, degree of saturation, and the substituent located on the third carbon (Fig. 7.2). Although it was originally believed that AHLs diffuse passively through cellular membranes, the long-chain molecules are actively transported via efflux and influx pumps
2.3. АГЛ-зависимая система Vibrio harveyi
Vibrio harveyi — грамотрицательная морская свободноживущая бактерия, способная к люминесценции. про экологию одно предложение.
Структура lux-оперона V.harveyi luxCDABEGH во многом схожа с V.fischeri: он тоже содержит гены субъединиц люциферазы (luxA и luxB) и гены ферментов, ответственных за синтез субстратов для биолюминесцентной реакции . Помимо этого, в lux-оперон V.harveyi входит ген luxH, чья функция пока остается неизвестной. Предполагается, что его продукт каким-то образом участвует в биосинтезе рибофлавина [Swartzman, Silverman, Meighen, с 7490]. Но главным отличием от lux-оперона V.fischeri является то, что ген АГЛ-синтазы (у V. fischeri это luxI) не входит в структуру оперона и является транс-регуляторным элементом.
Структуры lux-оперонов V.fischeri (A) и Vibrio harveyi (Б).
по диссеру коноплевой, с. 18
Несмотря на близкое родство, кворум-зависимая регуляция у V.harveyi сильно отличается от V.fischeri. На сегодняшний день у V.harveyi найдено три связанных системы чувства кворума, которые осуществляют совместную регуляцию транскрипции lux-оперона[]. Первая система
Аутоиндуктором первой кворум-зависимой системы регуляции транскрипции lux-оперона у Vibrio harveyi является типичный для грамотрицательных бактерий ацилгомосеринлактон (V. harveyi‘s AI-1 is N-(3-hydroxybutanoyl) homoserine lactone). Однако сама система регуляции устроена иначе и показывает большее сходство с системами чувства кворума грамположительных бактерий. АГЛ-синтазой
основана двух регуляторных компонентах: LuxS — АГЛ-синтазе, катализирующей реакцию синтеза …. из …. , и Lux
2.4 АИ-2 — системы
The QS regulon of V. harveyi is controlled by three separate QS systems. Each of these three systems has a distinct autoinducer synthase and a specific hybrid sensor histidine kinase protein. HAI-1 (harveyi AI-1) and AI-2 were discovered more than a decade ago (Bassler et al., 1994). V. harveyi‘s AI-1 is N-(3-hydroxybutanoyl) homoserine lactone (HSL), which is synthesized by LuxLM. This protein is not related to V. fischeri‘s LuxI but it performs a similar function (Federle and Bassler, 2003). N-(3-hydroxybutanoyl) homoserine lactone binds its specific sensor protein LuxN.
АИ-2 LuxS is required for the biosynthesis of AI-2, which is 3A-methyl-5,6-dihydro-furo(2,3-d)(1,3,2)dioxaborole-2,2,6,6A-tetraol. AI-2 is detected by a soluble periplasmic protein LuxP consequently leading to activation of a hybrid two-component sensor kinase response regulator protein, LuxQ.
АИ-3 The third QS system in V. harveyi was discovered more recently. The still uncharacterized autoinducer CAI-1 is synthesized by CqsA and detected by its cognate sensor CqsS. The Cqs system was first characterized in V. cholerae (Henke and Bassler, 2004a), which is where the name for this system comes from (cholerae quorum sensing).
все взаимосвязаны
The rest of the signaling cascade in V. harveyi is shared by all the systems (Fig. 7.5). LuxN, LuxQ, and CqsS dephosphorylate the shared phosphotransferase LuxU, which indirectly activates the response regulator LuxR.
The luminescence operon is activated by the LuxR protein, which requires a diffusible acylhomoserine lactone signal. The lux box, a 20 bp inverted repeat, is located in the luxI promoter region and is required for LuxR‐dependent induction of the luminescence genes. Using primer extension, we mapped the LuxR‐dependent transcrip https://onlinelibrary.wiley.com/doi/full/10.1046/j.1365-2958.1999.01261.x?sid=nlm%3Apubmed
Analysis of LuxR Regulon Gene Expression during Quorum Sensing in Vibrio fischeri https://jb.asm.org/content/189/11/4127
Cyclic AMP Receptor Protein Regulates Pheromone-Mediated Bioluminescence at Multiple Levels in Vibrio fischeri ES114
https://jb.asm.org/content/195/22/5051
http://genetika.ru/aaa%20Ucheniy%20Sovet/2016/Konopleva/2016_02_Konopleva_full_text.pdf диссер
QS controls activities that are unproductive when conducted by an individual cell, but become effective when performed collectively (Novick et al. 1995, Seed et al. 1995).
In the absence of the AI, most LuxR-type proteins do not fold correctly, which leads to proteolytic degradation or accumulation in inclusion bodies (Pinto and Winans 2009, Swem et al. 2009, Zhang et al. 2002), while LuxR:AI complexes are stable and can exert their cellular functions (Hussain et al. 2008, Piper et al. 1993).
План что дописать
- В историю — методы (?)
- Первый тип — что там с люксR (как регулируется), гомологи, двухкомпонентность
- Второй тип — харвеай, как устроена (гомологи?)
- третья — как устроена, гомологи, биологическая роль
An early study showed that a luminescent marine bacterium called Vibrio harveyi could sense a self-produced signal and also a signal or signals produced by other bacterial species to induce light production7. This phenomenon is considered to be a type of QS, where cells of many species in a mixed microbial community sense the general bacterial population density via a molecule termed Autoinducer-2 (AI-2)14,15
ЗАКЛЮЧЕНИЕ
Кворум-зависимые системы регуляции экспрессии встречаются у большинства известных грамотрицательных бактерий, где они регулируют самые разнообразные клеточные функции:
СПИСОК ЛИТЕРАТУРЫ
- Atkinson, Steve, and Paul Williams. “Quorum sensing and social networking in the microbial world.” Journal of the Royal Society, Interface 6,40 (2009): 959-78. doi:10.1098/rsif.2009.0203
- Mukherjee, S., Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nature Review Microbiology 17, 371–382 (2019). https://doi.org/10.1038/s41579-019-0186-5
- Pereira С.S, Thompson J.A., Xavier K.B., AI-2-mediated signalling in bacteria, FEMS Microbiology Reviews, Volume 37, Issue 2, March 2013, Pages 156–181, https://doi.org/10.1111/j.1574-6976.2012.00345.x
- Surette, M. G., Miller, M. B., Bassler, B. L. (1999). Quorum sensing in Escherichia coli, Salmonella typhimurium, and Vibrio harveyi: a new family of genes responsible for autoinducer production. Proceedings of the National Academy of Sciences of the United States of America, 96(4), 1639–1644. https://doi.org/10.1073/pnas.96.4.1639
- Swartzman E., Silverman M., Meighen E. A. The luxR gene product of Vibrio harveyi is a transcriptional activator of the lux promoter. Journal of Bacteriology. Nov. 1992, 174 (22) 7490-7493; doi: 10.1128/jb.174.22.7490-7493.1992
- Baldwin, T. O., Devine, J. H., Heckel, R. C., Lin, J. W., & Shadel, G. S. The complete nucleotide sequence of the lux regulon of Vibrio fischeri and the luxABN region of Photobacterium leiognathi and the mechanism of control of bacterial bioluminescence. Journal of bioluminescence and chemiluminescence, 4(1), 326–341 (1989). https://doi.org/10.1002/bio.1170040145
- Bassler, B. L., Wright, M., & Silverman, M. R. (1994). Multiple signalling systems controlling expression of luminescence in Vibrio harveyi: sequence and function of genes encoding a second sensory pathway. Molecular microbiology, 13(2), 273–286. https://doi.org/10.1111/j.1365-2958.1994.tb00422.x
- Bassler, B. L., Wright, M., Showalter, R. E., & Silverman, M. R. (1993). Intercellular signalling in Vibrio harveyi: sequence and function of genes regulating expression of luminescence. Molecular microbiology, 9(4), 773–786. https://doi.org/10.1111/j.1365-2958.1993.tb01737.x
- Eberhard, A., Burlingame, A. L., Eberhard, C., Kenyon, G. L., Nealson, K. H., & Oppenheimer, N. J. . Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry, 20(9), 2444–2449 (1981). https://doi.org/10.1021/bi00512a013
- Eberhard, A. Inhibition and activation of bacterial luciferase synthesis. Journal of bacteriology vol. 109, 3 (1972): 1101-1105.
- Engebrecht, J, M Silverman. Identification of genes and gene products necessary for bacterial bioluminescence. Proceedings of the National Academy of Sciences of the United States of America vol. 81,13 (1984): 4154-8. doi:10.1073/pnas.81.13.4154
- Engebrecht J, Nealson K, Silverman M. Bacterial bioluminescence: isolation and genetic analysis of functions from Vibrio fischeri. 1983 Mar;32(3):773-81 doi: 10.1016/0092-8674(83)90063-6
- Federle MJ, Bassler BL. Interspecies communication in bacteria. The Journal of clinical investigation vol. 112,9 (2003): 1291-9. doi:10.1172/JCI20195
- Fuqua WC, Winans SC, Greenberg EP. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. Journal of Bacteriology. 1994;176(2):269‐275. doi:10.1128/jb.176.2.269-275.1994
- Gambello, M. J., & Iglewski, B. H. (1991). Cloning and characterization of the Pseudomonas aeruginosa lasR gene, a transcriptional activator of elastase expression. Journal of bacteriology, 173(9), 3000–3009. https://doi.org/10.1128/jb.173.9.3000-3009.1991
- Greenberg, E.P., Hastings, J.W. & Ulitzur, S. Induction of luciferase synthesis in Beneckea harveyi by other marine bacteria. Archives of Microbiology. 120, 87–91 (1979). https://doi.org/10.1007/BF00409093
- Håvarstein, L. S., Coomaraswamy, G., & Morrison, D. A. (1995). An unmodified heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus pneumoniae. Proceedings of the National Academy of Sciences of the United States of America, 92(24), 11140–11144. https://doi.org/10.1073/pnas.92.24.11140
- Henke, J., Bassler, B. . Bacterial social engagements. Trends in cell biology, 14(11), 648–656 (2004). https://doi.org/10.1016/j.tcb.2004.09.012
- Higgins, D., Pomianek, M., Kraml, C. et al. The major Vibrio cholerae autoinducer and its role in virulence factor production. Nature 450, 883–886 (2007). https://doi.org/10.1038/nature06284
- Ji, G., Beavis, R. C., & Novick, R. P. (1995). Cell density control of staphylococcal virulence mediated by an octapeptide pheromone. Proceedings of the National Academy of Sciences of the United States of America, 92(26), 12055–12059. https://doi.org/10.1073/pnas.92.26.12055
- Kaplan, H. B., & Greenberg, E. P. Diffusion of autoinducer is involved in regulation of the Vibrio fischeri luminescence system. Journal of bacteriology, 163(3), (1985) 1210–1214.
- Kleerebezem, M et al. «Quorum sensing by peptide pheromones and two-component signal-transduction systems in Gram-positive bacteria.« Molecular microbiology vol. 24,5 (1997): 895-904. doi:10.1046/j.1365-2958.1997.4251782.x
- Manukhov I, Khrul’nova S, Baranova A, Zavilgelsky G. Comparative analysis of the lux operons in Aliivibrio logei KCh1 (a Kamchatka Isolate) and Aliivibrio salmonicida. Journal of bacteriology vol. 193,15 (2011): 3998-4001. doi:10.1128/JB.05320-11
- Miyamoto, C., Boylan, M., Cragg, L. and Meighen, E. (1989), Comparison of the lux systems in Vibrio harveyi and Vibrio fischeri. Biolumin. Chemilumin., 3: 193-199. doi:10.1002/bio.1170030406
- Miyamoto, C. M., Graham, A. F., & Meighen, E. A. (1988). Nucleotide sequence of the LuxC gene and the upstream DNA from the bioluminescent system of Vibrio harveyi. Nucleic acids research, 16(4), 1551–1562. https://doi.org/10.1093/nar/16.4.1551
- Nealson, K. H, and Hastings, J. W. Bacterial bioluminescence: its control and ecological significance. Microbiological reviews vol. 43,4 (1979): 496-518.
- Nealson KH, Platt T, Hastings JW. Cellular control of the synthesis and activity of the bacterial luminescent system. Journal of bacteriology vol. 104,1 (1970): 313-322.
- Papenfort, Kai, and Bonnie L Bassler. «Quorum sensing signal-response systems in Gram-negative bacteria.» Nature reviews. Microbiology vol. 14,9 (2016): 576-88. doi:10.1038/nrmicro.2016.89
- Passador, L., Cook, J. M., Gambello, M. J., Rust, L., Iglewski, B. H. (1993). Expression of Pseudomonas aeruginosa virulence genes requires cell-to-cell communication. Science (New York, N.Y.), 260(5111), 1127–1130. https://doi.org/10.1126/science.8493556
- Schaefer, AL et al. «Generation of cell-to-cell signals in quorum sensing: acyl homoserine lactone synthase activity of a purified Vibrio fischeri LuxI protein.» Proceedings of the National Academy of Sciences of the United States of America vol. 93,18 (1996): 9505-9. doi:10.1073/pnas.93.18.9505
- Schuster, M., & Greenberg, E. P. (2006). A network of networks: quorum-sensing gene regulation in Pseudomonas aeruginosa. International journal of medical microbiology: IJMM, 296(2-3), 73–81. https://doi.org/10.1016/j.ijmm.2006.01.036
- Swartzman, E., Silverman, M., & Meighen, E. A. (1992). The luxR gene product of Vibrio harveyi is a transcriptional activator of the lux promoter. Journal of bacteriology, 174(22), 7490–7493. https://doi.org/10.1128/jb.174.22.7490-7493.1992
- Tomasz, A. Control of the Competent State in Pneumococcus by a Hormone-Like Cell Product: An Example for a New Type of Regulatory Mechanism in Bacteria. Nature 208, 155–159 (1965). https://doi.org/10.1038/208155a0
- Waters, C. M., Bassler, B. L. (2005). Quorum sensing: cell-to-cell communication in bacteria. Annual review of cell and developmental biology, 21, 319–346. https://doi.org/10.1146/annurev.cellbio.21.012704.131001
- Лысак, В.В. Микробиология: учеб. пособие / В. В. Лысак. – Минск : БГУ, 2007.
или напишите нам прямо сейчас:
⚠️ Пожалуйста, пишите в MAX или заполните форму выше.
В России Telegram и WhatsApp блокируют - сообщения могут не дойти.
Прикрепленные файлы: |
|
|---|---|

|
Администрация сайта не рекомендует использовать бесплатные работы для сдачи преподавателю. Эти работы могут не пройти проверку на уникальность. Узнайте стоимость уникальной работы, заполните форму ниже: Узнать стоимость |

|
Скачать файлы: |
|
|
|